研究内容
私は樹木生態学を中心に、幾つかのテーマに沿って研究を行なってきました。
その中から、幾つかについて紹介したいと思います。
樹木の当年枝と繁殖生態
樹木は種によってさまざまな形態をしていますが、そのほとんどにおいて、枝や葉の茂った樹冠部と、垂直方向に伸びた幹部にわけることができます。

このうち樹冠部は、多くの場合、非常に複雑な形をしていて、中でどのようなことが起こっているのか、そしてどのような構造をしているのか把握することは困難です。しかし、その成長過程をよく見てみると、ある芽から枝が伸び、伸びた枝に再び芽ができて、その芽が芽吹いてまた枝を伸ばし、ということを繰り返していることがわかります。
1つの芽から1シーズンのうちに伸びた枝のことを「当年枝」と呼びますが、非常に複雑に見える樹木の樹冠部も、当年枝が繰り返し伸びていくことで出来ていっています。すなわち、樹冠部でどのようなことが起こっていて、どのような構造をしているのかは、当年枝を単位として観察、調査解析してゆくことで分かるでしょう。
ところで、樹木の活動のなかでも非常に興味深いのが繁殖活動です。樹木は動くことができませんので、繁殖によって種子を生産して散布することが、分布を拡大する唯一と言ってもいい手段です。ですので繁殖は非常に重要な活動であると言えます。

私が興味を持っているのは、当年枝を作って枝を伸ばすという活動と、繁殖という活動の両者をどのようにして上手く折り合いをつけているのか、という点です。一般的に言って、生物が使うことのできる資源の量は一定で、これを成長と繁殖という二つの活動の間で分け合います。すなわち片方が増えれば片方が減るという、「トレードオフ関係」にあります。枝をたくさん伸ばせば、それだけ多くの葉を茂らせて樹木としての成長は良くなりますが、そのぶん繁殖が減ると考えられます。逆に、たくさん繁殖をすれば、それだけ枝を伸ばすことができなくなってしまうと考えられます。では、当年枝を伸ばすという活動と、繁殖をするという活動の二つをどうやって折り合いをつけているのでしょうか。
私は、岐阜県高山市を調査地に、ヤマハンノキ(Alnus hirsuta var. sibirica)を材料として、当年枝の長さと、その当年枝が繁殖をするしないか、の関係を調べました。


その結果、明らかになったのは、ヤマハンノキには数センチに満たない短い当年枝から、100cmほどの長い当年枝までありますが、そのうち、花をつけている当年枝の長さは10cm以上40cm以下であるということです。
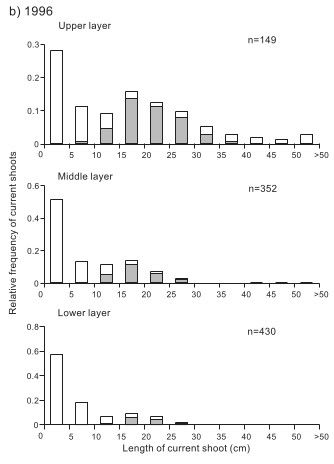
10cmよりも短い枝が花を着けないという結果は、このような短い枝では繁殖器官をつくるのに十分な資源を得ていないからだと考えられました。しかし、この考え方では、40cmよりも長い枝が花を着けないという結果を説明できません。
では、長い枝が花をつけ現象は、どういう時に起こるだろうかと考えて気づいたのが、ヤマハンノキのような高木の場合、繁殖を行う機会は一度だけではなく、何回もあるということです。
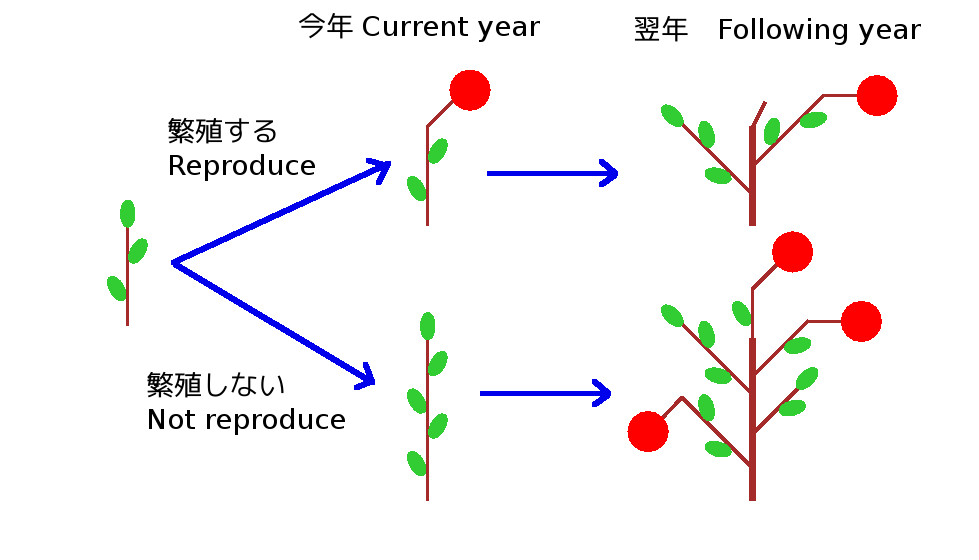
たとえば、ある当年枝があり、これが今年繁殖すると、一つ花をつけるとします。この枝が芽を2つ着けて翌年に2本の枝をつくり、うち1本が花を着けるとします(図の上側)。この場合2年間の花の合計数は1+1の合計2個となります。
一方で、同じ当年枝が今年繁殖しなければ、そのぶん枝を伸ばして芽を5つつけるとします。この芽が翌年に5本の枝になりうち3本が花を着けるとします(図の下側)。この場合、2年間の花の合計数は0+3の合計3個となります。もしこのような状況が成り立っているとすれば、この当年枝は、今年繁殖を行わないほうが2年間の合計としての繁殖量を増やすことができます。
私は、40cm以上の当年枝が繁殖を行わないことは、このような2年間の合計としての繁殖量を増加させるためではないかと考え、繁殖を行なった場合と行わなかった場合の繁殖量を推定してこれがヤマハンノキにおいて成り立つことを示しました。
これらの結果から私は、ヤマハンノキを構成する当年枝は、その長さに応じて、3つの機能グループに別れていると考えました。すなわち長さが10cm以下の、「維持型」のグループ、長さが40cm以上の「伸長型」のグループ、そして10cmから40cmの間の「繁殖型」のグループです。そして、このような機能の分化が、樹木個体の生涯の繁殖量を増加させる繁殖戦略であることを提唱しました。
余談:この論文が私の初めての国際誌での論文なのですが、ヤマハンノキの英名を”Japanese alder”と間違えてしまい、そのまま論文になってしまいました。その後も間違いに気付かず、学位論文まで間違いのまま通し、次の論文のとき査読者の方に指摘して頂いて初めて間違いに気づきました。正しくは”Siberian alder”です。この論文のタイトルを書くたび、恥ずかしさを思い出します。
- Hasegawa S. and Takeda H. (2001). Functional specialization of current shoots as a reproductive strategy in Japanese alder (Alnus hirsuta var. sibirica). Can. J. Bot. 79(1). 38-48.
以下、順次作成中
光合成産物の移動
樹木の当年枝は枝系を構成する形態上の基本単位の一つですが、これが生理的な単位となっているかは別問題です。
先のパートで当年枝の長さと繁殖の関係を示しているのですが、この議論には大きな穴が実はあって、「花は、その付いている枝にある葉で稼いだ光合成産物を使っている」ことを前提としてます。例えば、短い枝に花を着けて、この花に隣の長い枝で生産した光合成産物を運んで使うことが可能であったりすれば、話がだいぶ違ってきます(恐らくそんなことは無いであろうとは、様々な傍証がありましたが)。
繁殖器官は、その付いている枝から光合成産物を得ているかどうかを明らかにするため、炭素安定同位体で標識した光合成産物を与えて、その行方をたどる研究を行いました。

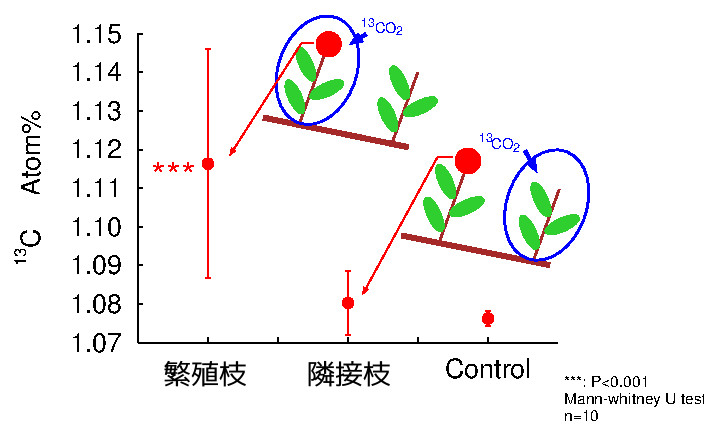
- Hasegawa, S., Koba, K., Tayasu, I., Takeda, H. and Haga, H.(2003). Carbon autonomy of reproductive shoots of Siberian alder (Alnus hirsuta var. sibirica). J. Plant Res. 116(3). 183-188.
- 長谷川成明(2003) 「樹体内の光合成産物の移動と樹木のモジュール性」 生物科学 54, 147-153.
樹形の進化
- Hasegawa, S. F., Shirota, T. and Kohyama, T. (2004) Simulation of evolutionary process of tree shape using cubic module model. Proceedings of 4th International Workshop on Functional-Structural Plant Models. 280.
景観生態学における推移行列の利用
- Takada, T., Miyamoto, A. and Hasegawa, S. F. (2010) Derivation of a yearly transition probability matrix for land-use dynamics and its applications. Landscape Ecology. 25 561-572.